Thermodynamik und energiereiche Verbindungen
Einer der universellen Energielieferanten ist das ATP. Es verfügt über drei Phosphorsäure-anhydridbindungen bei deren Hydrolyse Energie frei wird. Bei der Spaltung einer Phosphor-säureanhydridbindung
werden (-) 30,5 kJ/mol frei. Das ist gleichbedeutend mit der freien Energie nach Gibbs bei Standardbedingungen DG°’.
Der Ausdruck DG°´ bedeutet, dass die Reaktion bei pH 7, 25°C und in wässrigem Milieu stattgefunden hat. Die Gleichungen zur Berechnung lauten:
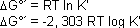
wobei R die allg. Gaskonstante ist, T die absolute Temperatur in ° Kelvin und K’ gleich dem Quotienten der Produkte über den Edukten.
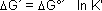
In der Natur kommen Reaktionen oft gekoppelt vor, denn so ist es möglich, dass Reaktionen mit positivem DG°´ ablaufen, die ohne Kopplung nicht ablaufen würden.
NAD+ (Nicotinamidadenindinucelotid) und NADP+ (Nicotinamidadenindinucleotidphosphat) besitzen wie der Name schon sagt zwei Basen: Adenin und Nicotin. Von den beiden ist Nicotin das aktive Zentrum des
Molküls. Es dient der Übertragung von Elektronen. Einen Vorstufe des NAD+ muss mit der Nahrung aufgenommen werden.
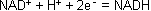
Übersicht zum Zuckermetabolismus 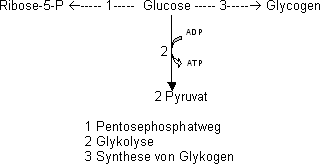
Alle diese Wege sind reversibel!
Die mit der Nahrung aufgenommene Stärke ist ein Glucosepolymer. Es wird im Darm wieder in Glucose zerlegt. Unser Körper braucht wenig Glucose um die Struktur zu erhalten, doch viel Glucose
wird unter ATP Gewinn zu CO2 abgebaut. In Pflanzen wird Glucose in Form von Stärke eingelagert, in Tieren in Form von Glykogen.
Die Glykolyse
Energiebilanz der Glykolyse
Man unterscheidet zwischen aerober und anaerober Glykolyse. Der Verlauf der Glykolyse ist zum grössten Teil identisch, doch am Schluss trennen sich die Wege.
aerob:
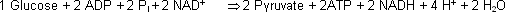
anaerob:
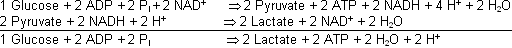
Erste Etappe der Glykolyse Die einzelnen Schritte der Glykolyse anhand der entsprechenden Enzyme:
Hexokinase Reaktion: 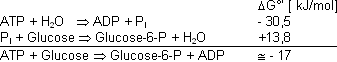
Die Hexokinase Reaktion ist eine gekoppelte Reaktion. Durch die Kopplung mit der Hydrolyse des ATPs wird die zweite Reaktion erst möglich.
Beachte: auch wenn eine Reaktion ein negatives DG°’ besitzt, muss diese Reaktion nicht unbedingt auch ablaufen, obwohl sie es vom energetischen Standpunkt her könnte!
Wichtig: Die Hexokinase Reaktion der Glykolyse ist eine irreversible Reaktion (grosses -DG der Gesamtreaktion). Für die Gluconeogenese (Umkehrung des Weges) wird ein anderes Enzym verwendet.
Das Enzym Hexokinase:
In der Hexokinase liegen die Bindungsstellen für Glucose und ATP sehr nahe beieinander. Die Kristallisation mit je einem gebundenem Substrat hat ergeben, dass zwei verschiedene Formen der Hexokinase
vorliegen. Das Enzym klappt von einer Form in die andere um beim Binden des entsprechenden Substrates. Beim Zuklappen des Enzym wird alles Wasser aus den aktiven Zentren entfernt. Dadurch wird erreicht,
dass nahezu immer das ATP für die Phosphorylierung der Glucose verwendet wird und nicht einfach mit Wasser reagiert.
Glucosephosphat-Isomerase:
Glucose-6-P › Fructose-6-P
Dies ist eine reversible Reaktion, bei der sozusagen keine Energie benötigt wird.
Phosphofructokinase:
Fructose-6-P + ATP › Fructose-1,6-bisphosphat + ADP
Eine zusätzliche P-Gruppe wird angefügt. Das Präfix „bis“ kennzeichnet, dass „zweimal“ phosphoryliert wurde. Diese Reaktion ist ATP abhängig und irreversibel.
Zweite Etappe der Glykolyse
Aldolase:
Die Aldolase schneidet die Fructose-1,6-bisphosphat in 2 Teile: Glycerinaldehyd-3-P + Dihy-droxyaceton-P. Diese beiden Verbindungen sind Isomere, die ohne grossen Energie-aufwand und ohne Coenzym von
der Triosephosphat-Isomerase ineinander umgewandelt werden › 2 Glycerinaldehyd-3-P, welche für den weiteren Verlauf der Glykolyse zur Verfügung stehen
Glycerinaldehyd-3-P-Dehydrogenase:
„Der geniale Schritt der Glykolyse“. Es ist der erste Schritt in Richtung Oxidation der Gluco-se. Die Dehydrogenase reduziert dabei NAD+ zu NADH (der Oxidationsstatus der Aldehyd-gruppe
des Glycerinaldehyd-3-P wird dabei zu der einer Carbonsäure ).
Glycerinaldehyd-3-P › 1-3-Bisphosphoglycerat
Phosphoglyceratkinase:
Das energiereiche P wird auf ein ADP übertragen. Erste Produktion von ATP. Es werden pro Glucose 2 ATP produziert, da zwei Moleküle 1,3-Bisphosphoglycerat pro Glucose gebildet wurden.
Der Name des Enzyms stammt von der Rückreaktion, der Gluconeogenese, her.
1-3-Bisphosphoglycerat › 3- Phosphoglycerat
Phosphoglyceratmutase:
Isomer-Reaktion: Verschiebung von P = Mutase
3- Phosphoglycerat › 2- Phosphoglycerat
Enolase:
Der Name Enolase beruht auf der Geschichte. Bei dieser Reaktion wird H2O eliminiert, was normalerweise von einer Dehydratase besorgt wird.
2-Phosphoglycerat › Phosphoenolpyruvat
Pyruvatkinase:
Der Name stammt wieder von der Rückreaktion her. Dies ist wieder eine irreversible Reaktion. Theoretisch ist zwar jede Reaktion reversibel, doch im menschlichen Körper stimmen die Bedingungen
für die Rückreaktionen oft nicht!
Phosphoenolpyruvat + ADP › Pyruvat + ATP (2. Produktion von ATP)
Im zweiten Teil der Glykolyse werden also pro Glucose 2x2 ATP gebildet. Abzüglich der zwei verbrauchten ATP im ersten Teil resultiert ein Nettogewinn von 2 ATP (+ 2 NADH, welche im aeroben Katabolismus
zur Bildung weiterer ATP führen – vgl. Krebs-Zyklus und oxidative Phosphorylierung !). Für Interessierte : aus einem Molekül Glucose können maximal (totaler Abbau zu CO2 und
H2O) 38 ATP gebildet werden.
Lactatdehydrogenase (anaerobe Glykolyse):
Pyruvat wird in Lactat umgewandelt, wenn kein O2 vorhanden ist. Die während der Gykolyse gebildeten NADH spenden die dazu nötigen Elektronen ( H- = H+ + 2 e-). Der Sinn der Lactatbildung
ist die Oxidierung der NADH zu NAD+, damit diese wieder für andere oxida-tive Reaktionen ( NAD+ als Oxidationsmittel ó wird selber reduziert) zur Verfügung stehen!
Energetische Aspekte der Glykolyse
ΔG°´ der aeroben Glykolyse beträgt » -84 kJ/mol. Während der ersten Etappe der Glykolyse werden 2 ATP verbraucht, in der zweiten Etappe jedoch 2x2 ATP gebildet.
Die Glycerinaldehyd-3-P-Dehydrogenase nimmt ein freies Pi und bildet eine energiereiche Säureanhydridbindung. Die Phosphorsäure trägt eine negative Ladung, welche Wasser anzieht. Wegen
des Wassermantels können phosphorylierte Substrate nicht mehr durch die Membran diffundieren und gehen der Zelle somit nicht mehr verloren.
Regulation der Glykolyse
Die Regulation der Glykolyse erfolgt auf dem Niveau der Phosphofructokinase (PFK). Die PFK besitzt mehrere allosterische Zentren für Citrat, ATP / AMP, ATP und Fructose-6-P. AMP und ATP bewirken
im gemeinsamen Zentrum eine andere Konformation.
Positive Effektoren sind: AMP und Fructose-2,6-Bisphosphat
Negative Effektoren sind: ATP und Citrat
ATP „diktiert“ der PFK, dass der Körper über genug Energie verfügt und kein neues ATP durch den Abbau von Glucose gebildet werden muss.
Da ein ständiger Wechsel von AMP ‹› ADP ‹› ATP im Körper stattfindet, gibt auch AMP Auskunft über den Energievorrat.
Die Gluconeogenese
Die Gluconeogenese, also die Neubildung von Glucose, findet hauptsächlich in der Leber statt, doch auch die Niere ist daran beteiligt. Ausgangsstoffe für die Gluconeogenese sind Lactat oder
Alanin, welche in Pyruvat umgewandelt werden oder andere AS die direkt in Oxalacetat umgewandelt werden und so in die Gluconeogenese einsteigen. Es gibt somit mehrere „Einstiegsorte“ für
die Gluconeogenese.
Gluconeogenese ist nicht gleich Umkehrung der Glykolyse
Die Glucose-6-Phosphatase ersetzt in der Leber die Hexokinase und bildet aus Glucose-6-Phosphat Glucose. Die Hexokinase ist aus energetischen Gründen nicht reversibel.
Bei der Hydrolyse von GTP entsteht Wärme, die verloren geht.
Die Pyruvatcarboxylase braucht Biotin (Vitamin H) um aus Pyruvat Oxalacetat herstellen zu können. Das Biotin muss über die Nahrung aufgenommen werden. Das CO2 wird über Biotin an der
Pyruvatcarboxylase gebunden. ATP und Pyurvat werden ebenfalls an dieses Enyzm gebunden, jedoch an einer anderen Bindungsstelle. Sind alle Substrate gebunden, faltet sich das Enzym ab und ermöglicht
so die Übertragung des CO2 auf das Pyruvat.
Phosphoenolpyurvatcarboxykinase: sobald CO2 vom Oxalacetat abgespalten wird, entsteht ein sehr reaktives Enolat, welches die Phosphat Gruppe des GTP angreift und sie abspaltet. Die Umwandlung von Pyruvat
erfordert also die Hydrolyse von einem ATP und einem GTP › sehr energieaufwendig !
Die Gluconeogenese kann z.T. auch in den Mitochondrien stattfinden. Allerdings kann Oxalacetat nicht durch die Mitochondrienmembran transportiert werden. Darum wird es in Malat umgewandelt, weil
für Malat Transporter zur Verfügung stehen. Im Zytosol wird aus Malat wieder Oxalacetat aufgebaut. Aus Oxalacetat wird dann Phosphoenolpyruvat (PEP) synthetisiert.
Ein wenig PEP wir auch in den Mitochondrien hergestellt und kann via Transporter ins Zytosol gelangen.
Bilanz der Gluconeogenese
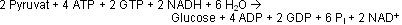
Die Gluconeogenese verbraucht neben ATP auch GTP!
Bei ATP Überschuss:
ATP + GDP ‹› GTP + ADP
Die Gluconeogenese ist keine Umkehrung der Glycolyse: es werden 6 ATP-Equivalente + 2 NADH verbraucht! Die Umkehr der Glycolyse würde nur 2 ATP + 2 NADH benötigen, doch die Umkehrung ist
energetisch in der Zelle nicht möglich.
Die Gluconeogenese verbraucht Energie. Herrscht in den Leberzellen Energiemangel, wird keine Glucose synthetisiert, denn dann brauchen die Leberzellen die Energie zum Überleben.
Regulation der Gluconeogenese
Das regulierte Enzym der Gluconeogenese ist die Fructose-1,6-bisphosphatase:

Positive Effektoren der Gluconeogenese wirken auf die Glycolyse hemmend, währen negative Effektoren aktivierend wirken. Die Gluconeogenese und die Glycolyse werden somit reziprok reguliert. Es
ist sehr wichtig, dass nicht beide Wege gleichzeitg in einer Zelle ablaufen können, da sonst Energie „sinnlos“ verbraucht würde.
Pentosephosphatweg (auch Hexosemonophosphatweg)
Aus Glucose wird Ribose hergestellt, welche ein wichtiges Strukturmolekül ist und z.B. für die Nukleinsären (DNA und RNA) benötigt wird.
Man kennt zwei Wege um Glucose in Ribose umzuwandeln, einen oxidativen und einen nicht oxidativen Weg.
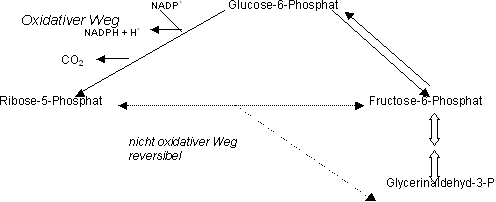
Braucht eine Zelle nur NADPH, wird die Glucose über Ribose zu Gycerinaldehyd-3-P weiterabgebaut und nachher via Fructose-6-P zu Glucose aufgebaut (oxidativer Weg).
Benötigt die Zelle aber Ribose und kein NADPH erfolgt der Abbau der Glucose über den nicht oxidativen Weg, d.h. über Fructose-6-Phosphat zu Ribose.
Der oxidative Pentoseweg
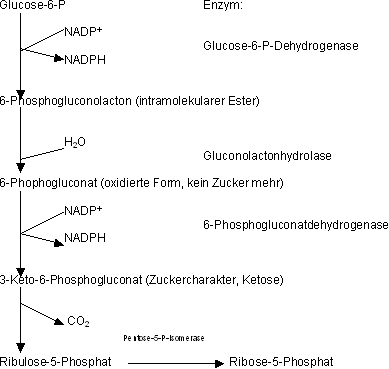
Bilanz des oxidativen Weges
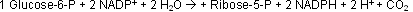
Das zentrale am oxidativen Weg ist die Synthese von NADPH, welches in vielen Enzymen, die e- benötigen, zu NADP+ oxidiert wird.
Der nicht oxidative Pentoseweg
Aus Ribose5-P wird dabei Fructose-6-P gebildet. Die einzelnen Umwandlungsschritte sind: 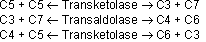
Die Transketolase transferiert C2 und die Transaldolase C3. Ist der C2/C3 Donator eine Ketose, wird er zu einer Aldose. Ist der Donator jedoch eine Aldose, wird er zur Ketose.
Bilanz des nichtoxidativen Pentoseweges
3 Ribose-5-P ‹› 2 Fructose-6-P + Glycerinaldehyd-3-P
Die Regulation erfolgt durch die Konzentration der vorhandenen Substrate.
Der Unterschied zwischen NADH und NADPH
Das Verhältnis der Konzentrationen in den Zellen:
NADPH : NADP+ = 100 In den Zellen finden wir beinahe nur die reduzierte Form NADPH
NADH : NAD+ = 0,001 Die Zellen oxidieren NADH sofort zu NAD+
Für Oxidationen (e.g. Glykolyse, Krebs-Zyklus) verwenden Zellen also NAD+, während sie für Reduktionen (e.g. Fettsäuresynthese) NADPH benötigen.
Regulation der Durchflussmenge
Die vorliegenden Substrate, also NADPH und Ribose, bestimmen darüber, über welchen Weg Glucose zu Ribose umgewandelt wird und in welcher Menge dies geschieht. Verfügt die Zelle über
genügend Substrate, muss keine Glucose abgebaut werden.
Glucose-6-P-Dehydrogenase Deficency (Fabismus)
Die menschlichen Erythrozyten haben keine eigene Proteinsynthese (kein Kern mehr !), doch auch sie benötigen NADPH um Schäden, die durch O2 entstanden sind, zu beheben. Das dafür verantwortliche
redoxaktive Molekül ist das Glutathion, ein Tripeptid mit Cystein in der Mitte. Die aktive Gruppe des Glutation ist die Thiolgruppe (SH) des Cysteins.
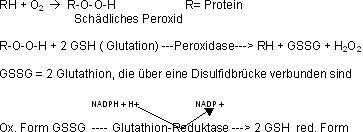
Fehlt die Glutathion-Reduktase, sammeln sich oxidierte Proteine und Lipide in den Erythrozyten an und bewirken den vorzeitigen Tod. Kann nun wegen fehlender Glucose-6-P-Dehydrogenase nicht genügend
NADPH hergestellt werden, kann das Glutathion nach der Enfernung der schädlichen Peroxide aus den Proteinen nicht mehr reduziert werden, was zu einen frühzeitigen Tod der Erythrozyten durch
Peroxide führt.
Bei einer genetischen Krankheit, der Glucose-6-P-Dehydrogenase-Deficency, ist genau dies der Fall und die Erythrozyten sterben vorzeitig. Unter normalen Bedingungen ist es verkraftbar, dass die Erythrozyten
schon nach etwa 90 Tagen sterben und nicht erst nach 120 Tagen. Durch den Genuss einer bestimmten Bohnensorte wird jedoch die kritsche Grenze für den Abbau der Erythrozyten erhöht und es werden
mehr Erythrozyten als gewöhnlich abgebaut. Es kommt zur hämolytischen Anämie.
Malariaerreger können in Zellen mit Glucose-6-P-Dehydrogenase-Deficency nicht gut überleben. Darum ist diese Krankheit in Malariagebieten ein Selektionsvorteil. Da das Gen für diese
Krankheit auf dem X- Chromosom liegt, sind nur die heterozygoten Frauen resistent gegen Malaria. Wieso nicht auch die Männer? Sind alle Erythrozyten von Glucose-6-P-Dehydrogenase-Deficiency betroffen
kann sich der Malariaerreger anpassen und auch dort leben. Bei den heterozygoten Frauen sind jedoch nur die Hälfte von den Erythrozyten von Glucose-6-P-Dehydrogenase-Deficency betroffen und machen
es dem Malariaerreger somit schwer sich anzupassen.
|
